
Revisiting the Fisher King
/
I know, I know, my last post promised a series on reconstructing Acrocanthosaurus in multiple views - bear with me, as this is actually part of that series. Remember that both animals have stuff sticking up on their backs, so I want to be able to compare and contrast those elongated neural spines...and how those differences should impact reconstructions of the animals. But to do that I had to update this skeletal, as new information had rendered the older one no longer tenable.

Besides, Spinosaurus is cool! For one, it's the only dinosaur in the Jurassic Park series to tangle with a T. rex and emerge victorious (no matter how unlikely that outcome was). It's probably the longest theropod we know of, and may have been the heaviest as well. Yet counter-intuitively it shows specialization for piscivory (fish-eating)...maybe in JP3 the spinosaur mistook the T. rex for a really large lung-fish?
Tongue firmly out of cheek now, Spinosaurus has lit up imaginations partially due to its size, but also because there was so much you had to imagine to try and reconstruct the animal. Until the last decade or two it was sort of a theropod Rorschach test where you could project any sort of oversized monster theropod onto its scant (and now lost) remains. This brings a thrilling "Sherlock Holmes" quality when trying to imagine the living animal, but for most of the last century serious attempts to reconstruct Spinosaurus have been more frustrating than titillating.
Darren Naish has an excellent write up of the history (and tragedy) of of the type specimen of Spinosaurus, which I won't duplicate here. The long and short of it is that WWII claimed the fossils as another victim of the conflict. The already-meager remains lost, paleontologists were stuck with the original description and some somewhat uninspired sketches as the only link to the past.
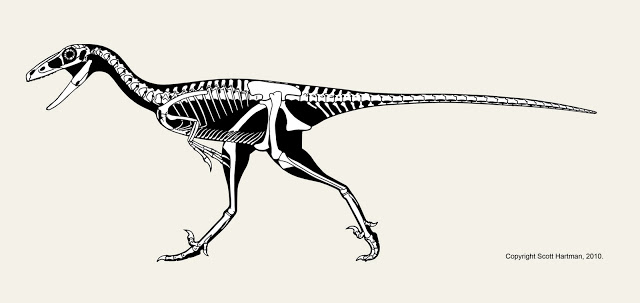
A series of fortunate events occurred in the latter half of the 20th century that allowed for a more accurate interpretation of Spinosaurus to emerge. For one, other spinosaurids were found. Baryonyx from the U.K., and Nigerian Suchomimus, started to paint a more complete picture of what these animals were like. They had bizarrely long snouts that seemed to resemble a gharial as much as a traditional theropod. Suchomimus even had a smaller version of the enlarged neural spines on the back:

The amusingly-named Irritator from South America further clarified the relationships and anatomy of spinosaurids. But the real breakthrough was the re-discovery of several photographic plates of the original material. While Spinosaurus wasn't the most complete specimen, having photographs at least made it possible to ensure that what was found is incorporated accurately into a reconstruction.
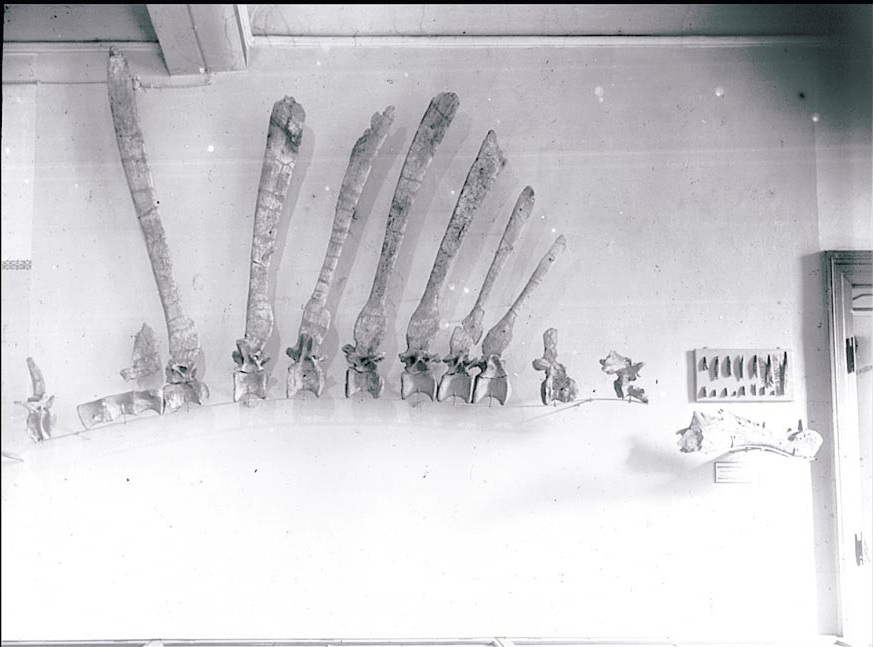
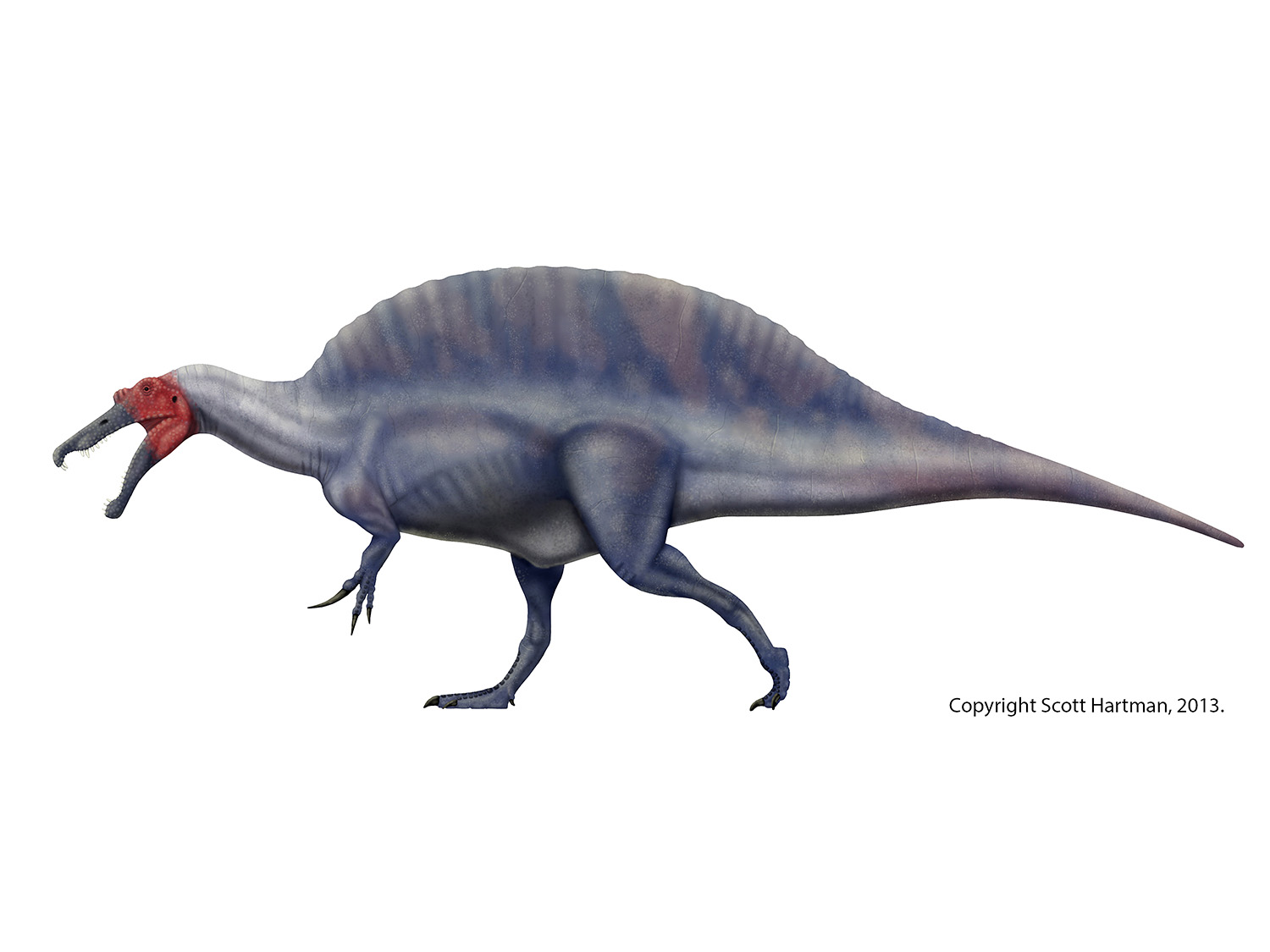
Among other details, the image also shows what had been the basis of attempts to restore the shape of the elongate sail or hump on the back: Stromer's original interpretation for the position of the elongated neural spines. In particular, notice that the tallest one is set directly in front of the sacrum here, while the only associated tail vertebra (at the far left of the picture) has a very short spine. That has lead most people to infer that the spine started quickly after the neck, grew to ridiculous heights over the pelvis, and then quickly dropped off again. Indeed, this is the interpretation that I used in my first attempt, and has been widely seen in such disparate and reputable scientific endeavors as Jurassic Park 3, the Carnegie Collection of "museum quality replicas", and Greg Paul's reconstruction in his Princeton Field Guide to Dinosaurs.
And they're in excellent company (whereby I arbitrarily define myself as "excellent company"). I had been concerned with Stromer's original interpretation for the placement of the tallest neural spinse - no vertebral body (centrum) was preserved, but the change in the angle of the spine seemed pretty extreme compared to the previous dorsals, especially right in front of the sacrum. My solution was to assume it was a sacral neural spine. This largely preserved the traditional appearance of the "sail", but provided a bit of breathing room for the change in orientation.

Luckily for us, Andre Cau and Jamie Headden were busy mulling over this specific issue, and came to a much more likely conclusion, that the backward-oriented neural spine was actually an anterior caudal. Looking at a host of dinosaurs with elongate neural spines, they noted that in general you never seen backward-canted spines in front of the hips, you always see them after it. There is a bit more detail to the argument (which I encourage you to read on their blogs), but in essence they make a very compelling case.
And so it was back to the virtual drawing board. I made some other corrections from my previous attempt - there had been some scaling issues with the neck vertebrae that had given my reconstruction a thinner Baryonyx-like profile in the neck. Also, it appears that the necks of these animal don't have as much of the traditional theropod S-curve, so that was changed as well (although I still don't buy the extreme hang-dog look that Greg Paul has started to restore his spinosaurs with). The results are a stockier animal, with a more elongate sail (or hump):

Looking at the rigorous reconstruction, it's clear that there's still quite a bit of uncertainty in the skeleton, although not all of the missing parts are created equal. Much of the pelvic girdle is known from Irritator, as is the back of the skull. Also, some unpublished specimens shed light on this, even if they aren't documented well enough to be official parts of the reconstruction. Still, there's a bit of ambiguity about the exact limb proportions, the length of the tail, and the exact shape of the sail.
Speaking of which, how should those tall neural spines be restored by artists doing life reconstructions? Is it a sail, was it supporting a hump of tissue like a bison, or was it simply a muscular ridge? We'll get back to that subject in a bit, after looking at Acrocanthosaurus.
Until then, best wishes to one and all for a wonderful 2012!


.jpg)

























{kind=link}