
Gauging stance in "wide-gauge" sauropods
/
In 1999 Jeff Wilson and Matt Carrano published an excellent paper addressing the phenomena of "wide-gauge" sauropod trackways. For years researchers had been working to explain why sauropod trackways seemed to come in two very different flavors - some of them were very closely spaced...so much so that they would actually overlap on the midline of the track. Other sauropod tracks seemed to show animals walking with their feet spread much further apart.
What were paleontologists to make of this?
One explanation was that the trackways were made by the same type of sauropods that were engaging in different behaviors. In other words, perhaps sometimes a sauropod would walk with its legs close in, while at other times it would use a wide-gauge stance.
Wilson & Carrano proposed that instead the trackways were made by sauropods with different skeletal adaptations. They mustered quite a few lines of evidence, but perhaps the best was that there was a group of sauropods - titanosaurs - that in fact had a much wider pelvis than other sauropods. The paper created a framework for later workers to use when attempting to correlate track makers with fossilized trackways, and is generally a towering success.
But I did want to take issue with one figure of the paper - one that pops up repeatedly at SVP. It is figure 5, demonstrating their interpretation of hing leg stance:

That's Camarasaurus on the left and Opisthocoelicaudia on the right. The clever reader may have already surmised from the title of this post that I think the animal on the right has its legs spread too far apart. But I have a larger issue: both animals have their legs spread much too far apart.
Remember that narrow-gauged trackways actually have their feet fall so close together that they frequently overlap along the midline. There's no way even sauropod "A" could make those tracks in the stance as figured. And this is why I'm bringing this up, because animals generally don't walk around with their legs acting as perfectly vertical beams. If you spend time watching large animals walk away from you, you'd see something like this:

(Elephant image from here, rhino image from here.)
People also move like this, with our vertical limbs generally sloping in toward the midline when we walk or run. There are probably several reasons for this (including mechanical efficiency) but for our purposes here let's just setting on the fact that it happens. Large, straight-limbed graviportal animals tend to walk with the limbs angled inward, not down (and certainly not angled out).
And the trackways also demonstrate this. If you place place sauropods over the actual trackways in question, you end up with a stance more like this:
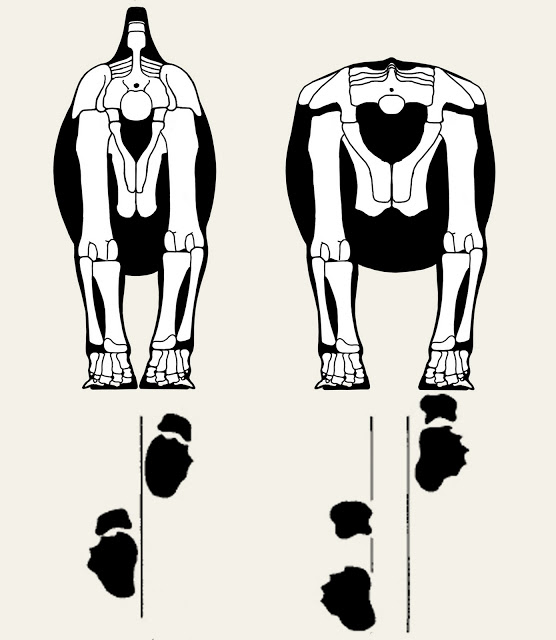
In this case I've put a diplodocid (Supersaurus) on the left, while the animal on the right is scaled to the pelvic dimensions of Opisthocoelicaudia as seen in the original paper. Both animals have the hind legs mostly vertical but gently sloping inward.
This is not to say that sauropods never adopted a pose with their legs spread out a bit; Wilson & Carrano point out that titanosaurs have adaptations that may have allowed them to evert their hind limbs more effectively. They suggest that titanosaurs may have done so when rearing up, or during other activities that require greater stability.
I don't take issue with that, and those sorts of differences in the legs and pelvis may make it possible to tease out further behavioral differences between sauropod groups. But when walking around in their day to day lives both the footprints and modern analogs make a strong case that the limbs should be vertical, and if anything sloping in towards the midline rather than spread away from it.
Reference:
Wilson, J. A, & Carrano, M. T. 1999. Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology, 25(2), pp. 252-267.

























{kind=link}